|
|
Journal of Advanced Veterinary Research Volume 9, Issue 3, 2019, Pages: 123-127 www.advetresearch.com |
|


|
Anatomical Studies on the Nasal Plane of Camels Revealing Feasible Functions |
|
|
|
Eman A. Eshrah |
|
|
|
Department of Anatomy and Embryology, Faculty of Veterinary Medicine, Benha University, Egypt (eman.eshrah01@fvtm.bu.edu.eg) |
|
|
|
Received: 1 June 2019; Accepted: 2 July 2019 |
|
|
|
Abstract |
|
The nasal plane (rhinarium) in camels is an important specialized integumentary structure, however, little is known about its structure and function. Accordingly, the first aim of this study was to describe the microscopic anatomy of the dermis. The second aim was to determine the anatomical communications of the nasal plane, in relation to the feasible functions. The microscopic study revealed a characteristic dermal structure. While, the gross anatomical study revealed important anatomical communications between the nasal plane and the vomeronasal organ openings. Together, the discussed data indicating that the nasal plane in dromedaries may play a role in delivering stimuli to the vomeronasal organ. |
|
Keywords: Camel, Dromedary, Nose; Rhinarium, Nasal plane, Dermis. |
|
|
|
Introduction |
|
|
|
The nasal plane (rhinarium) is a newly described integumentary structure in camels (Eshrah, 2017). Because of its very small size and the unusual location that veiled by the philtrum edges, it has been hidden from the eyes of the observers for a long time. Thus, little is known about its structure and function. The anatomy of camel rhinarium was different from that of the other domestic ruminants. For example, in caprine and ovine, the nasal plane covers the whole medial nasal wing and extends in bovine to cover also the upper lip (Nickel et al., 1979). However, in camels, it was confined to a small area covering the philtrum and the medial nasal angles. Conversely, the camel nasal plane was greatly similar to the type found in the proboscis-bearing mammals, in particular, Moose (Alces alces) (Clifford, 2003). For the above mentioned reasons, studying the nasal plane in camels will add important information to the field of comparative anatomy. As a part of the integumentary system, the nasal plane is an indicator of body health (Dellmann and Brown,1987). Additionally, it plays a significant role in identification of animal individuality (Maala and Lalusis, 2000). However, there is no available information regarding its functional correlations. The fine structure and the anatomical relations are important keys to understand organ functions. There are some important nearby structures, including, openings of the vomeronasal duct system and incisive papilla. These structures are essential for detection of liquid phase molecules (LPM) found in food, urine and genital secretions. In turn, LPM molecules require initial treatment with mucous secretions (Nagashima and Touhara, 2010; Wood, 2012). The nasal plane is permanently covered with mucus secretions. Additionally, it’s the first part touch the animal food as well as LPM carrying secretions. This raise the question, why not the nasal plane plays a role in delivering stimuli to the VNO? this motivates the author to investigate the anatomical communications between the nasal plane and this organ. Additionally, studying the dermal structure is important to clarify the glandular content, which together with the underlying and nearby structures, will help understanding the possible functions of the nasal plane in this species. This also may help explain why its anatomical plan greatly differs from that of the other ruminants. For these, the first aim of the current work was to clarify the microscopic anatomy of the dermis in the camel rhinarium. The second aim was to determine the feasible functions of the nasal plane in this species. |
|
|
|
Materials and methods |
|
Animals The present study was carried out on twelve heads of apparently healthy adult (3-5 years) male camels. The heads were obtained immediately after slaughter from Toukh abattoir. Any lesions or congenital abnormalities were considered excluding criteria. This study was approved by the Institutional Animal Care and Use Committee of Benha University, Egypt. Study design The current study underwent two steps: the first step was a microscopic study, the second was a gross examination to help understanding the functional anatomy of the nasal plane. First step: Five heads were used for the microscopic study. Second step: Seven heads were used to examine the nearby mucosal openings and to define the feasible communications between theses openings and the nasal plane. Light microscopy This study was undertaken on the glabrous skin of the philtrum (median labial groove) and the medial nasal angles. The selected samples were obtained immediately after slaughter. These samples were fixed in 10% neutral formalin, dehydrated in ascending grades of alcohol, cleared in xylene and embedded in paraffin wax. Sections of 3 µm in thickness were obtained and stained by Harris heamatoxylin and Eosin (H&E) for general histological architectural examination according to Bancroft and Stevens (1997). |
|
|
|
Results |
|
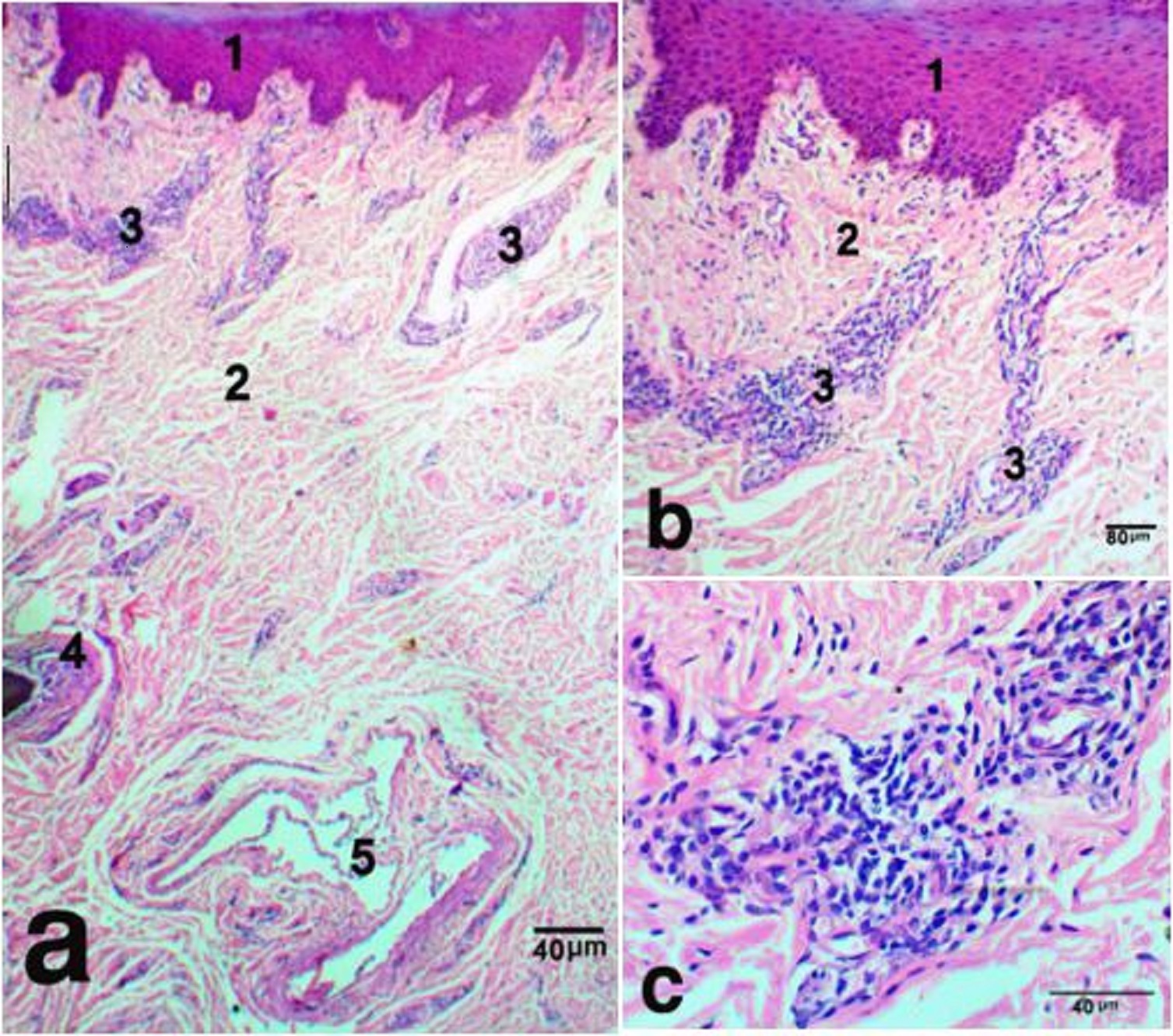
Microscopic study The dermis of the nasal plane was formed of two continues layers, the superficial papillary and the deep reticular layers. The superficial layer was immediately adjacent to the epidermis and conformed to the contours of its basal layer, forming finger like extensions, the dermal papillae. These papillae were tightly fitted to their equivalent the epidermal pegs. They decreased in height toward the hairy side. The papillae were mostly of the vascular type. They contained blood capillaries and infiltrated with immune-responsive cells, such as, lymphocytes, neutrophiles, and macrophages. However, some papillae, which were located near the junction with hairy skin, contained the excretory portion of sweat glands (Fig. 1a,b).
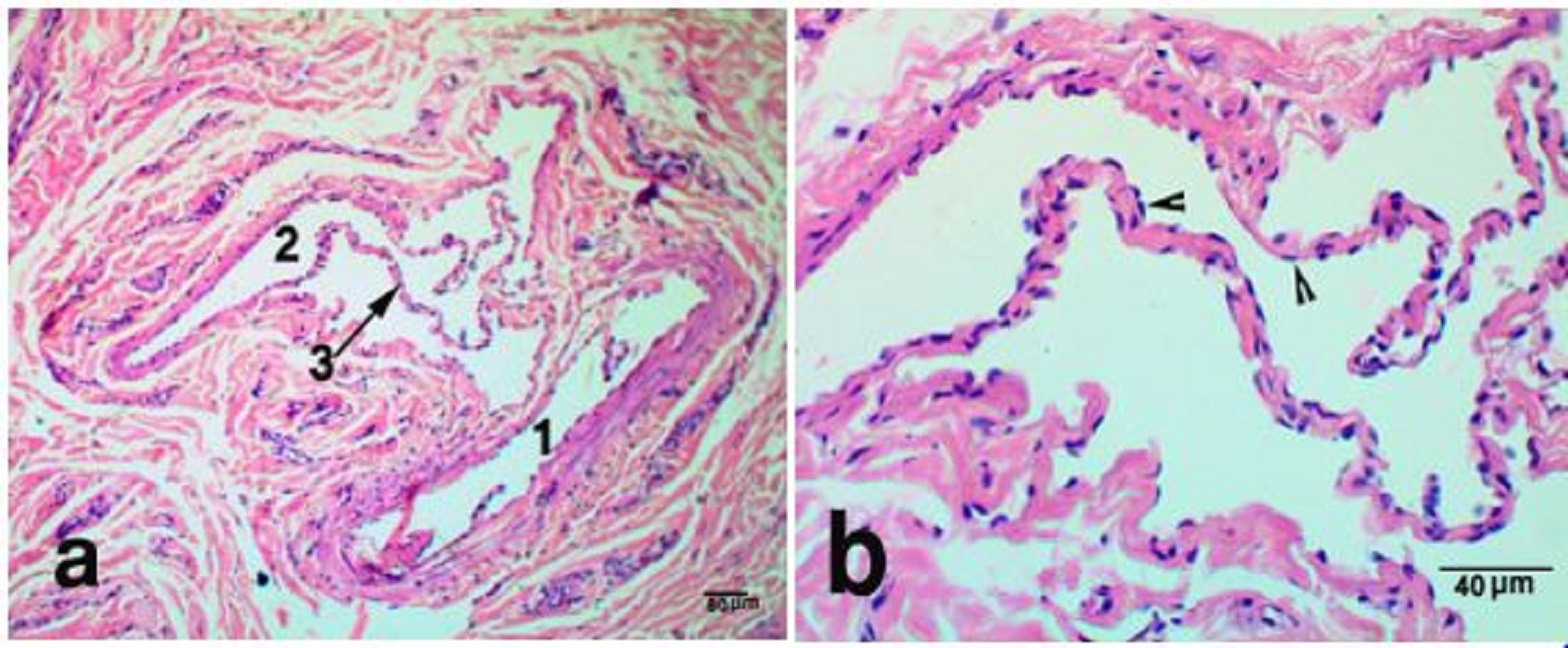
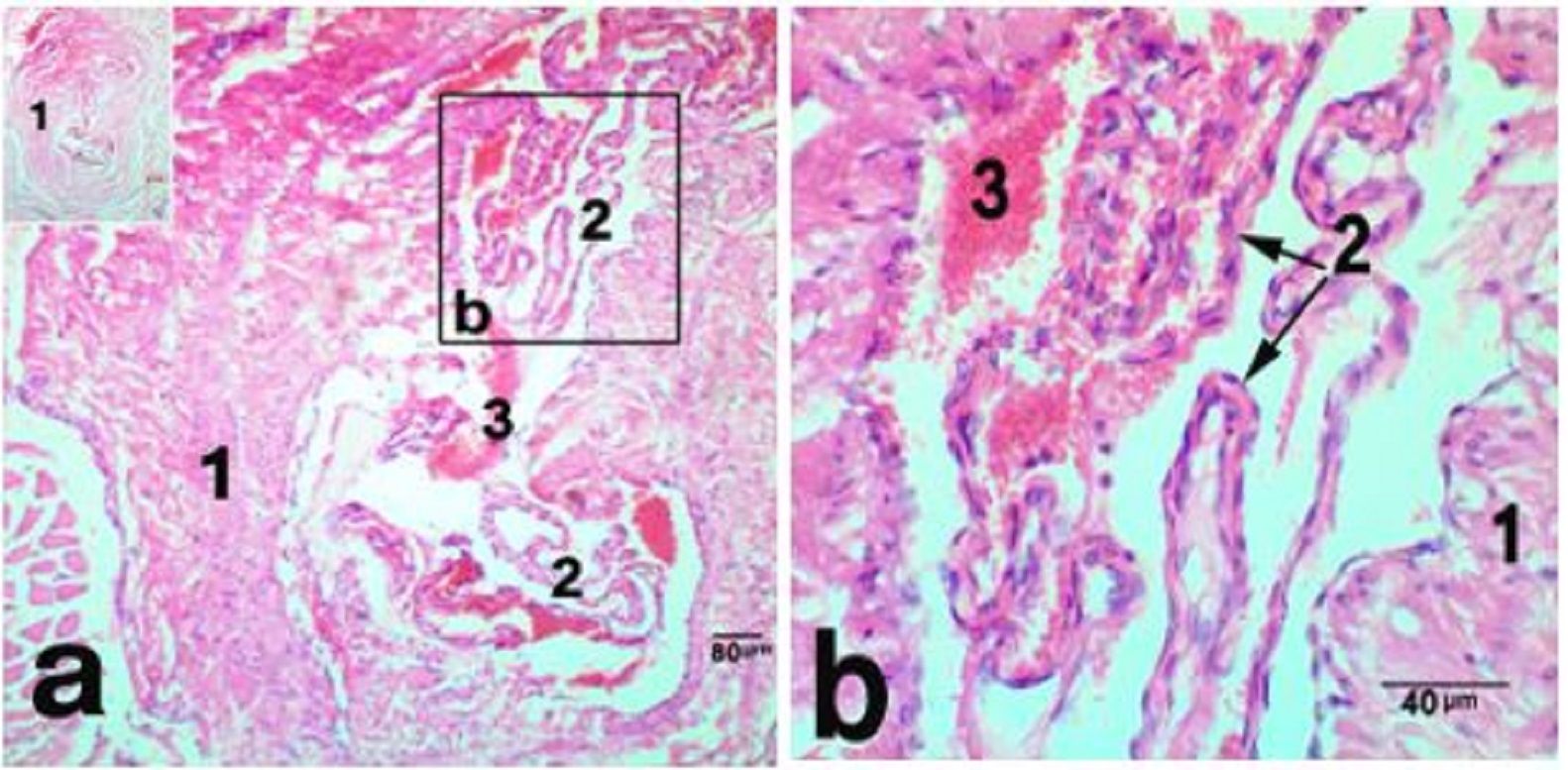
Fig. 1. The nasal plane in camels, an area adjoining hairy skin, H&E stain, X 40 (a), sweat glands X 100 (b) and X400 (c), showing, epidermis (1), dermis (2), sweat glands (3), hair follicle (4), simple vascular anastomosis (5). The reticular dermis was formed of dense irregular connective tissue, which continued distally with muscular subcutis. Atypically, the dermal core was devoid of sebaceous, sweat or sero-mucus glands, except for a narrow zone, adjoining the hairy skin. This area had numerous sweat glands that opened onto the surface epithelium through sweat pores (Fig. 1a,b). They were winding and highly coiled tubular merocrine glands (Fig. 1b,c). Deeply in this area, two types of Arterio-venous anastomoses (AVAs) were observed. They were simple and complex anastomoses in the form of vascular bridge and glomus body, respectively. The vascular bridge anastomosis was consisted of a single endothelial layer surrounded by myoepithelial cells (Fig. 2 a,b). The glomus body was consisted of multiple layers of endothelial cells that surrounded by a well-innervated connective tissue capsule (Fig. 3 a,b). The vascular bridge was located in the superficial reticular dermis, while the glomus body found deeper in the dermis or subcutis.
Fig. 2. Vascular bridge AVNs, in the dermis of nasal plane, H&E stain X 100 (a) and X 400 (b) showing, arterial side (1), venous side (2), the vascular bridge (3) a single endothelial layer forming the vascular bridge (arrow heads).
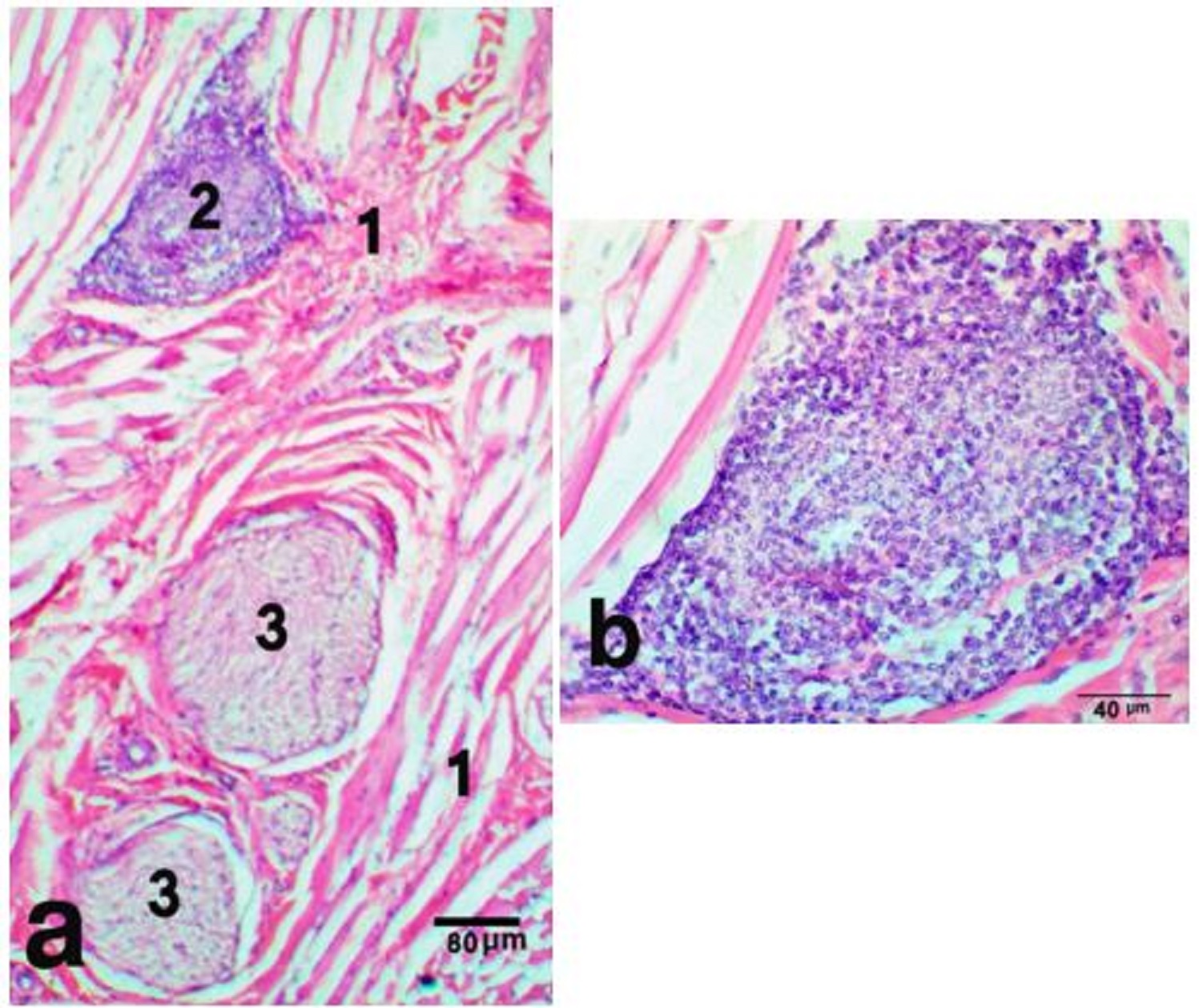
Fig. 3. Glomus body in the nasal plane, H&E stain X 100 (a) and Multiple endothelial layers forming the bulk of the glomus, H&E stain X 400 (b) showing, connective tissue capsule (1), endothelial layers (2), blood cells (3). Subcutis was blended with the underlying labial musculature, and entirely formed of skeletal muscle fibers of orbicularis oris muscle. It was a complex muscle of no clear points of origin and insertion. It was formed of interlacing muscular fibers that intermingled with those of nearby muscles. These muscle fibers were contained numerous proprioceptive muscle spindles (Fig. 4 a,b). The subcutis was attached the skin of the nasal plane to the lip mucous membrane. Groups of labial, serous glands were noticed parallel to the distal part of the philtrum.
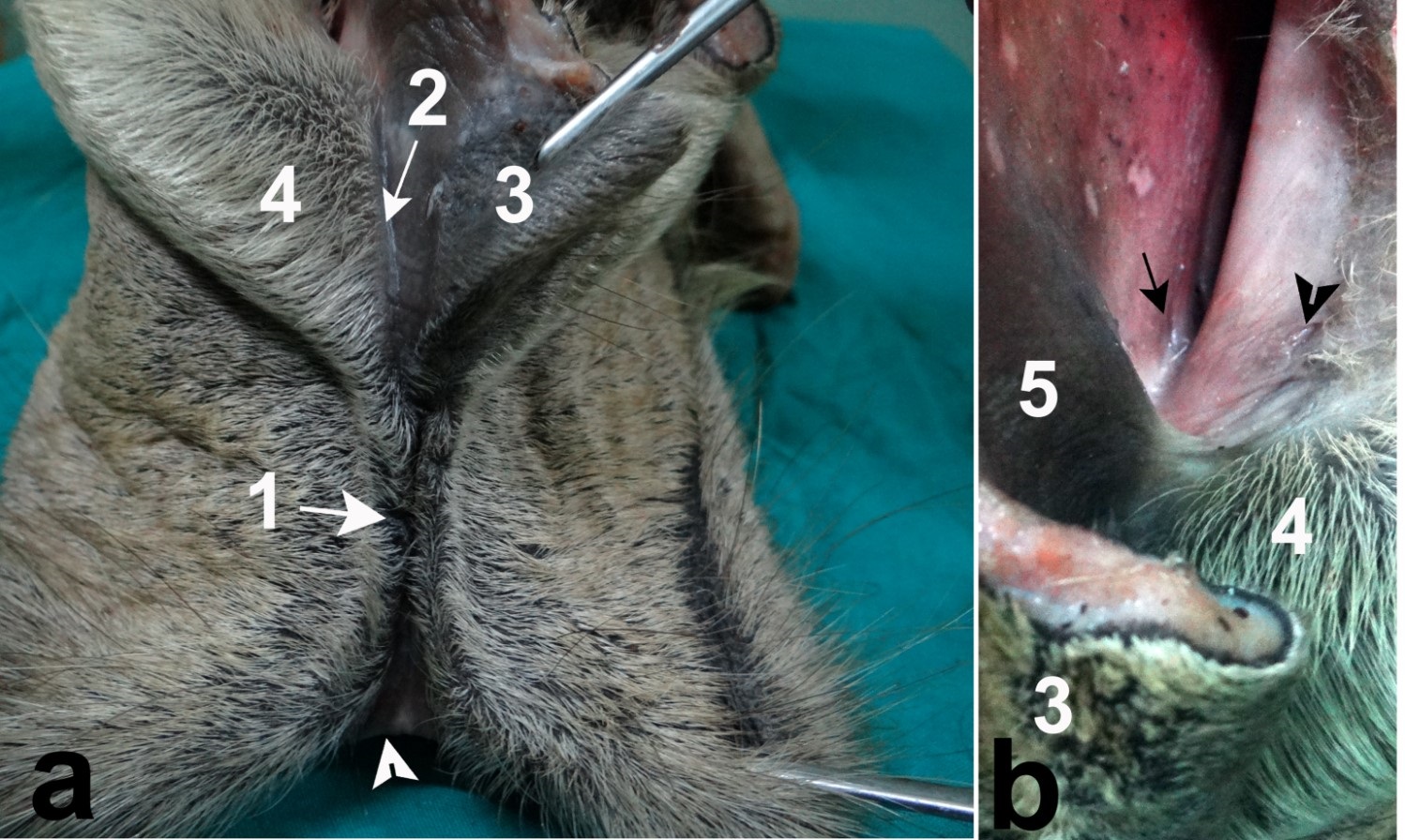
Fig. 4. Muscle spindle in the subcutis of nasal plane, H&E stain X 100 (a), notice, the nerve axons wrap around the muscle fibers forming nuclear bag, H&E stain X 400 (b), skeletal muscle fibers (1), nuclear bag fibers (2), nerve axon (3). Grosss anatomical study The perinasal part of the nasal plane was continued medially into the nasal vestibule as a hairless groove. This groove was mainly paraseptal, extended in close proximity and parallel to the nasal septum. It continued caudally to end near the common opening of the vomeronasal organ (VNO) and incisive duct (Fig. 5a). Retraction of the upper lip can easily sweep the mucus covering the nasal plane through this paraseptal groove into the VNO opening. The openings of the lateral nasal diverticulum and the nasolacrimal duct were located at the muco-cutaneous junction (Fig. 5b). They were in contact with the leading edge of the paraseptal groove, where this groove formed a steeper angle that can allow receiving secretions of the lateral nasal diverticulum and the nasolacrimal duct.
Fig. 5. Anatomical Communications of the nasal plane (a), nearby glandular openings (b), distal end of nasal plane (white arrow head), edges of philtrum (1), paraseptal groove (2), medial nasal wing (3), lateral nasal wing (4), muscular part of nasal septum (5), opening of the lateral nasal diverticulum ( black arrow head), opening of the nasolacrimal duct (black arrow). |
|
|
|
Discussion |
|
The present study revealed the microscopic structure of the dermis in the nasal plane of dromedary camels. The dermis was atypically devoid of nasolabial and sebaceous glands. Additionally, the wide capillaries and large blood vessels were not observed. Sweat glands and arterio-venous anastomosis were confined to a narrow zone adjoining the hairy skin of the lips. The subcutis was formed entirely of skeletal muscle fibers rich in muscle spindles proprioceptors. Conversely, in the types found in other ruminants, the mucus and sebaceous glands are typically present, the large thick-walled blood vessels are abundant and the hypodermis is formed entirely of adipose tissue (Dellmann and Brown,1987; Nickel et al, 1979; Aughey and Frye, 2001). Based on the anatomical results it was hypothesized that the nasal plane in camels may function as an associated structure of the vomeronasal system. Trap and dissolve odore molecules in the covering film of secretions. Then, retraction of the upper lip can sweep the dissolved molecules into the VNO opening via the paraseptal groove. Because of the attachment of the muscular subcutis to the upper lip musculature, the nasal plane able to retract in conjunction with the upper lip and enable the nasal plane to deliver stimuli into the VNO. Supporting this hypothesis is that camels lack VNO oral communication. The incisive duct, which is the oro-nasal passage that delivers stimuli to the VNO (Døving and Trotier, 1998) ends blindly in the hard palate (Karimi et al., 2014). Thus, the camel rhinarium may work as a functional substitute. Generally, the mammalian rhinarium is covered by a permanent layer of mucus, supplied by the underlying dermal glands (Nickel et al., 1979; Dellmann and Brown,1987). It has been confirmed by observations in living camels that the rhinarium was constantly covered by a transparent viscous film of secretions (Eshrah, 2017). The role of mucus secretions is essential for olfaction. It dissolves scents and separate their components chemically, this make them available for detection by the neuroreceptors (Nagashima and Touhara, 2010; Wood 2012). Lack of glandular contents was normally present in canine rhinarium, in which the surface mucus substituted by the lacrimal secretions. Although the role of lacrimal secretions is not clear. It may trap and dissolve odor molecules, which cannot dissolve in mucus secretions. Similarly, the current study suggested extradermal sources for the surface secretions. These sources include, the nasolacrimal ducts and the lateral nasal diverticuli. Supporting this idea, the presence of two paraseptal groove as steep passages between their openings and the nasal plane. The lateral nasal diverticuli or sacs are unusual cylindrical pouches, on either side of camel face, which open into the nasal vestibule (Arnautovic and Abdalla, 1969; Smuts and Bezuidenhout, 1987; Metwally et al., 2019). It produces mucous secretions that moisten the inhaled dry air of the desert and may also act as reservoirs (Magid and Razag, 1975). The present study suggested that the main source of mucus covering the nasal plane may be the lateral nasal sacs. They open immediately at the caudal limit of paraseptal groove, which can sweep their secretions to the nasal plane continuously during rest. Active supply is also feasible, as the sac lies between the facial muscles and consequently compresses when these muscles work (Eshra and Badawy, 2014). All the facial muscles around the nostrils are inserted without tendinoues attachment into the orbicularis oris (Eshra and Badawy, 2014). During retraction of upper lip, as in flehmen response, the orbicularis oris can indirectly compress the sacs by grasping their surrounding muscles. This may provide fresh and copious secretions that help trap larger amount of airborne odor molecules. Sweat glands are not uncommon in the skin of camels (Taha and Abdallah, 1980; Fath-Elbab et al., 2017). They are essential for the thermal regulation in withstanding climate extreme in the desert (Schmidt-Nielsen et al., 1957; Fath Elbab et al., 2017). Sweat glands and AVN had special distribution in the nasal plane. They present only in the area adjoining hairy skin, leaving the dermal core devoid of glands and large blood vessels. This distribution, suggesting a thermoregulatory mechanism that provides a stable adherent layer of surface mucus. Stability of the temperature and humidity of mucus is crucial for its function (Nagashima and Touhara, 2010; Wood, 2012). It directly, affects its ability to trap or dissolve the odorant molecules. |
|
|
|
Conclusion |
|
The present microscopic and anatomical studies support the hypothesis that the camel rhinarium may help delivering dissolved stimuli into the VNO opening. This may functionally substitute for the lack of VNO oral communication in this species. However, further studies are still recommended to verify this hypothesis. |
|
|
|
Funding sources Author covered full cost for this work. |
|
|
|
Conflict of Interests |
|
No conflict of interests have been declared. |
|
References |
|
Arnautovic, I., Abdalla, O., 1969. Unusual blind sac on the face of the one humped camel. Acta Anatomica 73, 272-277. Aughey, E., Frye, F.L., 2001. Comparative Veterinary Histology with Clinical Correlates. 1st Ed. Manson publishing, The Veterinary Press. Spain, pp. 263-267. Bancroft, J.D., Stevens, A.A., 1997. Theories and Practice of Histological Technique, 4th, Chuchill Living Stone, Livingstone Edinburgh, London, New York. Clifford, A.B., 2003. Narial novelty in mammals: case studies and rules of construction. M.Sc. Faculty of the college of arts and science, Ohio University, U.S. A. Dellmann, D., Brown, E.M., 1987. Textbook of Veterinary Histology, 3rd Ed., Lea and Febiger, Philadelphia, U.S.A. Døving, K., Trotier, A., 1998. Structure and function of the vomeronasal organ. Journal of Experimental Biology 201, 2913–2925. Eshra, E.A., Badawy, A.M., 2014. Peculiarities of the camel and sheep narial musculature in relation to the clinical value and the mechanism of narial closure. Indian Journal of Veterinary Anatomy 26, 1, 10-13. Eshrah, A.E., 2017. The Camel Rhinarium: A study revealing the presence of the nasal plane in dromedary camel (Camelus dromedaries), with special reference to its epidermal structure. Anat. Histol. Embryol. 46, 65-72. Fath El-Bab, M.R., Abou-Elhamd, A.S., Abd-Elkareem, M., 2017. How the structure of the sweat glands of camel symphonizes their reliable function. J. Anim. Health Prod. 5, 19-23. Karimi, H., Mansoori, R., Ale Hashem, Ardalani G., Sadrkhanloo R., Hayatgheibi H., 2014. Structure of Vomeronasal Organ (Jacobson organ) in Male Camelus Domesticus Var. dromedaris persica. Anat. Histol. Embryol.43, 423–428 Maala, C. P., Lalusis, A.D.L., 2000. The anatomy of muzzle (Planum nasolabiale) and muzzle prints (Nasolabiogram) classification in Holstein Friesian Cattle and their crosses.Philipp. J. Vet. Med. 37, 1–8. Magid, A.M., Razaq, A.I., 1975. Relationship and possible function of the nasal sacs and glands of the one-humped camel, Camelus dromedarius. Acta Anat. (Basel) 91, 3, 423-8. Metwally, M.A., Hussieni, H.B., Kassab, A.A., Eshra, E.A., 2019. Comparative Anatomy of the Nasal Cavity in Buffaloes, Camels and Donkeys. J. Adv. Vet. Res. 9, 69-75. Nagashima, A., Touhara, K. 2010. Enzymatic Conversion of Odorants in Nasal Mucus Affects Olfactory Glomerular Activation Patterns and Odor Perception. The Journal of Neuroscience 30, 16391–16398. Nickel, R., Shummer, A., Seiferle, E., 1979. Viscera of domestic animals, 2nd, Verlag Paul Parey. Berlin, Hamburg. Schmidt-Nielsen, K., Schmidt-Nielsen, B., Jarnum, S.A., Houpt, T.R., 1957. Body temperature of the camel and its relation to water economy. Am. J. Physiol. 188, 103–112. Smuts, M.S., Bezuidenhout, A.J., 1987. Anatomy of the Dromedary.1st Ed. Clarendon Press, Oxford. U.S.A. Taha, A.A.M., Abdalla, A.B., 1980. Light and electron microscopy of the sweat glands of the dromedary camel. Acta Vet. Brno 49, 31-35 Wood, A.W., 2012. Physiology, biophysics and biochemical engineering. CRC press Taylor and Francis group New York, pp. 475-501. |
|
|