|
|
Journal of Advanced Veterinary Research Volume 10, Issue 2, 2020, Pages: 88-95 www.advetresearch.com |
|


|
Pathological Investigation on Natural Infection by Pasteurella Multocida in Goats |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Aziza Amin |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Pathology Department, Faculty of Veterinary Medicine, Benha University, 13736, Egypt. (aziza.amin@fvtm.bu.edu.eg) |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Received: 21 February 2020; Accepted: 28 March 2020 |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Abstract |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Pasteurellosis caused by Pasteurella multocida is, one of the most important infectious diseases of goats resulting in huge economical loss, not been previously reported in Egyptian native goats. This study was carried out to diagnose P. multocida depending on clinical examination, necropsy, bacterial culture and histopathology as well as pasteurella antigens detection by immunohistochemistry and then to compare the results of immunohistochemistry (IHC) with other traditional diagnostic methods. Thus, a clinicopathological examination was done on twenty Egyptian native goats (1-24-month old) showed clinical manifestations of nasal discharges, frothy salivation, weakness, in-appetence and eventual deaths. Necropsy consistently displayed severe fibrinous pleuritis, pneumonic patches in many lobes with focal hemorrhage and abundance of sero-sanguinous fluid along the thoracic regions. Hemorrhage with scattered yellowish necrotizing lesions was observed on the liver and spleen. Histopathology of the lungs revealed fibrinous and suppurative bronchopneumonia with massive distension of the pulmonary alveoli with inflammatory cells, red blood cells and fibrin. Various pathological alterations were also observed in the liver, heart, kidney, spleen and intestine. The isolated bacteria were identified as P. multocida. PCR and IHC positive reaction of P. multocida was detected in the lung, liver, kidney, heart, spleen and intestine. It could be concluded that the gross and histopathological lesions may be helpful in initial diagnosis of pasteurellosis in goat but it has to be complemented by other diagnostic tests. IHC method evidenced to be a consistent, accurate, and fast technique to incorporate these traditional pasteurellosis diagnostic approaches. |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Keywords: Pasteurellosis, Goat, Histopathology, Immunohistochemistry |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Introduction |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Pasteurella organisms are β-hemolytic, gram-negative, aerobic, non-motile, non-spore forming commensal coccobacilli in the family Pasteurellaceae. Pasteurella multocida is a normal commensal in the mucosal surfaces of the upper respiratory tract, gastrointestinal tract and genital tract of animals and in any stress state or infections with other agents, P. multocida proliferates and resulting in variety of diseases in different animal species (Dabo et al., 2007). P. multocida is one of the most relevant cause of pneumonia in cattle, sheep, and goats. It has been distinguished as one of the main bovine respiratory disease complex associated pathogens, resulting in loss of productivity and high mortality. Generally, pneumonia is conducting economical losses in ruminant industry all over the world (Ackermann and Brogden, 2000). The most popular strain of the respiratory pasteurellosis isolated from sheep and goat is P. multocida as it has been reported as a cause of a variety of lesions such as mild exudative to necrotizing pneumonia as well as exudation into bronchi and bronchioles. Furthermore, P. multocida causes also septicemia and systemic pasteurellosis in sheep at age less than two months. Dyspnea, listlessness, anorexia and sudden death were the most common characteristics of the systemic form of P. multocida infection in young sheep. However, if more than 20% of the lung was damaged, chronic breathing problems can occur, including reduced lung capacity and weight gain (Watson and Davis, 2002). In certain cases, such as antibiotic treatment, frozen tissue, autolytic material and other conditions, is often difficult to identify pasteurella with bacteriological methods. However, the immunohistochemical (IHC) methods have removed several problems related to other detection methods for pasteurella antigens on fixed, paraffin embedded tissues (Haziroglu et al. 1996). Consequently, the current research was aimed to diagnose goat pasteurellosis using post-mortem and the microscopical examination with Immunohistochemistry staning technique and to compare the effectiveness of immunohistochemical (IHC) techniques with conventional diagnostic methods. |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Materials and methods |
|||||||||||||||||||||||||||||||||||
|
Animals A total of 20 native breed goats (1-24 months old) in farms at Kaluobia governorate with case history of respiratory manifestations were admitted for post-mortem examination to the animal clinic at the Faculty of Veterinary Medicine, Benha University Egypt, during the period of 2018-2019, Collection of Samples The samples were collected from different organs for bacterial isolation, polymerase chain reaction, histopathological examination and immunohistochemistry. The current work was conducted following the approval of the Research Committee and the Institutional Animal Ethics Committee for animal care and experimentation at the Faculty of Veterinary Medicine, Benha University. Bacteriological examination Both blood agar containing 5% sheep blood and MacConkey agar media were used for bacterial isolation from heart blood, lung, liver, kidney, spleen and intestine. The plates were incubated at 37°C for 18-24 hours, then examined for bacterial growth. The detected colonies were checked according to Quinn, et al. (2011) for identification of the morphology of Pasteurella multocida like colony. In addition, the suspected colony was stained by gram stain following Ozyildiz et al. (2013). Smears from heart blood and the impression smears from Lungs were prepared, and stained with gram stain to detect bipolarity. Polymerase chain reaction (PCR) Specimens from lung, heart, kidney, liver, spleen and intestine were collected then preserved at -20°C for PCR analysis. DNA extraction from these samples was performed using the QIAmp® extraction kit (Qiagen, Vienna, Austria) following the manufacturer’s instructions. The used primers were developed from the bacterial genome of P. multocida. The forward primer had the sequences 5'-AATGTTTGCGATAGTCCGTAAGA-3', while the sequences of the reverse one was 5'-ATTTGGCGCCATATCACAGTCG-3' (Accession No. AF067175). Then, in a T gradient thermocycler (Biometra, Goettingen, Germany) following the protocol of Gautam et al. (2004), the resulting mixture was exposed to a particular thermal profile as follows: an initial denaturation cycle at 94 °C for 30 seconds; followed by 30 cycles at 94°C for 50 seconds, 30 seconds at 55°C and at 72°C for 1 minute; followed by one final extension cycle of 72° C for 10 minutes. A molecular marker of 100 bp DNA ladder was used on 1.5% agarose gel to evaluate the amplified products (10 μl). Visualization of amplicons with ethidium bromide by using a UV trans-illuminator with a wavelength of 590 nm and positive reactions were determined by size (540 bp) in length. Pathological examination After internal examination was completed in necropsied animals, samples from lungs, liver, kidney, heart, spleen and intestine were taken and fixed in 10% neutral buffered formalin then the fixed tissue were processed, embedded in paraffin wax then sectioned (4 µm) and stained with hematoxylin and eosin (H&E) according to Bancroft and Gamble (2007) for demonstration of any histopathological alterations in the examined samples. Immunohistochemistry (IHC) Samples from lungs, liver, kidney, heart, spleen and intestine were used for detection of the bacterial antigen by IHC staining technique according to Narita et al. (2000). Post deparaffinization and rehydration, of 3 μm-thick tissue sections, antigen retrieval by heating the slides in citrate buffer (pH 6.0) in a microwave oven was performed. These slides were then washed in phosphate buffer saline for 20 min. The activity of endogenous peroxidase has been blocked with 0.5% hydrogen peroxide (H2O2) in methanol for 30 minutes. The sections were then incubated with 1:10 dilution of a normal goat serum (Sigma, G9023) mixed with 2% bovine serum albumin for 45 minutes at room temperature in a humidified chamber, followed by an overnight incubation with hyperimmune serum prepared in rabbit for P. multocida antibody with dilution of 1:4000 in PBS. Then sections were extensively washed with PBS three times. To visualize the primary antibody, the sections were incubated for 30-minutes at room temperature by an induction of 1:500 dilution of biotinylated anti-rabbit IGG secondary antibody (Vector Laboratories, Burlingame, USA), followed by the Vectastain ABC Kit (Vector Laboratories, Burlingame, USA) for thirty minutes. The reaction has been demonstrated with a freshly prepared 3, 3-diaminobenzidene tetrahydrochloride (DAB) Substrate Kit for peroxidase (Vector Laboratories, Burlingame, USA). Then counter staining was done by using Gill’s hematoxylin stain (Merck, Germany). Exclusion of primary antibodies has been applied for negative control. Further, the intensity of IHC reaction was completed on the basis of antigen positive cells and classified as mild, moderate, and severe. Autoclaved scissors and disposable forceps were used for sampling, to avoid cross contamination during necropsy examination. |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Results |
|||||||||||||||||||||||||||||||||||
|
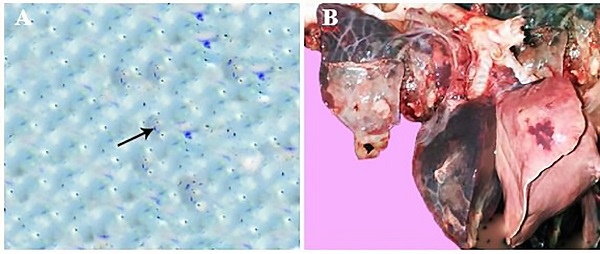
Clinical findings The main clinical signs observed on the affected animals were anorexia, cough, mucoid nasal discharge, depression and death. Gross examination The most common gross lesions demonstrated in the lungs of the most cases were pneumonic patches with dark red areas and consolidation of many lobes with firm consistency as well as congestion, focal hemorrhages, fibrinous pleuritis and emphysematous patches with frothy exudate oozing out on cut section were observed (Fig. 1B). Additionally, marked congestion, hemorrhage with scattered yellowish necrotizing lesions was observed on the hepatic surface, spleen and the intestine of most examined cases. Bacteriological findings P. multocida was bacteriologically isolated from lung lesion, heart blood, liver, kidney, spleen and intestine. On blood agar plate colonies appeared as small colonies (1-2 mm in diameter). P. multocida appeared as gram-negative, non–spore-forming coccobacilli with bipolar staining features. Additionally, blood smears from heart blood and impressions from pulmonary exudate revealed number of bipolar organisms indicating the presence of Pasteurella spp (Fig. 1A).
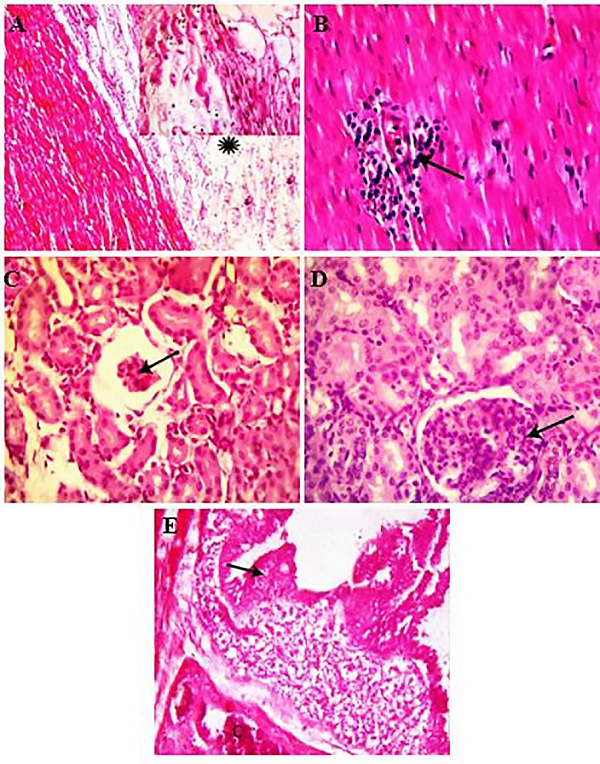
Fig. 1. (A) Bipolarity of P. multocida in pulmonary exudate stained with gram stain (arrow, x1000) (B) pneumonic patches with dark red patches, consolidation of many lobes and firm consistency with focal hemorrhages, notice also fibrinous pleuritis. Polymerase chain reaction P. multocida was identified by PCR in numerous tissues (Table 1). PCR reaction revealed positive amplification of specific bacterial DNA fragment in the lungs, heart, liver, spleen, kidney and intestine. The maximum rate of positive PCR reaction (95%) were observed in pulmonary tissues. Furthermore, positive signals were also noticed in 80% of the processed heart samples followed by hepatic tissue as positive PCR signals were demonstrated in 60% of the examined liver samples. 40% of the splenic tissues showed positive PCR signals. Meanwhile, the lowest percentage of positive PCR reaction was recorded in the kidney (30%) followed by the intestine (15%). Histopathological findings The main pathological alterations associated with P. multocida were represented mainly by fibrinous, suppurative and fibrinopurulent bronchopneumonia. Congestion and thrombosis of pulmonary blood vessels, hemorrhage in the inter-alveolar space with distention of pulmonary alveoli with inflammatory cells and erythrocytes was detected (Fig. 2A). Additionally, the bronchioles were filled with purulent exudates containing poly-morphonuclear inflammatory and necrotic cells, with peri-bronchial connective tissue proliferation admixed with mononuclear leukocytic cellular infiltration as well as distension of alveoli and bronchiole with inflammatory exudate admixed with different inflammatory cells, including mainly neutrophils, lymphocytes and macrophages in association with erythrocytes. (Fig. 2B,C). Meanwhile, fibrinous bronchopneumonia was also demonstrated in which pulmonary alveoli was packed with fibrin and some inflammatory cells (Fig. 2D). On the other side, the bronchioles and alveoli in other examined cases were found distended with fibrino-purulent exudate characterized by the loosely filled cellular infiltrates containing mononuclear and polymorphonuclear cells, interspersed with fibrinous exudates into the alveoli with thickening of alveolar wall with fibrin (Fig. 2E,F). Necrotizing bronchopneumonia associated with degeneration and necrosis of the bronchial epithelium and distention of respiratory bronchioles and alveoli with leukocytic cellular infiltration and eosinophilic proteinaceous edematous substances (Fig. 3A). Furthermore, necro-suppurative bronchopneumonia in which necrosis of the epithelial cell lining of the bronchioles with the presence of eosinophilic inflammatory exudate admixed with inflammatory cells mainly neutrophils in its lumen as well as focal areas of coagulative necrosis infiltrated with leukocytes was detected in the pulmonary tissue (Fig. 3B). In addition to this, multifocal hemorrhagic areas were demonstrated in the interstitial spaces in association with proliferation of interstitial tissue with mononuclear leukocytic cellular infiltrations was also detected. Interestingly, chronic active bronchopneumonia characterized by Thickening of alveolar septa with fibrin and inflammatory cells in association with multiple hemorrhagic areas in the pulmonary tissue were also observed (Fig. 3C). Furthermore, hyalinization of the alveolar content with marked thickening of the inter-lobular septa was also demonstrated (Fig. 3D,E). Obvious thickening of the pleura with varying degrees of leukocytic cellular infiltration were noticed (Fig. 3F).
Fig. 2. H&E stained section of lung obtained from goat naturally infected with P. multocida, showing (A) thrombosis of pulmonary blood vessels (asterisk) with distension of alveoli with mononuclear leukocytes and erythrocytes (arrow, x200), (B) the bronchioles filled with purulent exudates containing polymorphonuclear inflammatory cells (asterisk) with peri-bronchial connective tissue proliferation admixed with mononuclear leukocyte (arrow, x400), (C) distension of the bronchiole with inflammatory exudate admixed with different inflammatory cells, necrotic substances and fibrin (x 200), (D) pulmonary alveoli was packed with mixture of fibrin and inflammatory cells (x200), (E) distension of the bronchioles with fibrino-purulent exudate (x200), (F) the alveoli were loosely packed with cellular infiltrates containing mononuclear and polymorphonuclear cells, interspersed with fibrinous exudates (arrow) with thickening of alveolar wall with fibrin (x400).
Fig. 3. H&E stained section of lung obtained from goat naturally infected with P. multocida, showing (A), degeneration and necrosis of the bronchial epithelium and distention of respiratory bronchioles and alveoli with leukocytic cellular infiltration and eosinophilic proteinaceous edematous substances (x200) (B), necrosis of the lining epithelium of the bronchioles (arrow) with packing of its lumen with eosinophilic inflammatory exudate admixed with inflammatory cells mainly neutrophils (asterisk) as well as focal areas of coagulaive necrosis infiltrated with leukocytes in the pulmonary parenchyma (zigzag arrow, x200), (C) distension of alveoli with eosinophilic edematous substances mixed with different inflammatory cells and fibrin, notice also, multiple hemorrhagic areas in the pulmonary tissue (x 200), (D), hyalinization of the alveolar content with mild leukocytic cellular infiltration. Notice also thickening of alveolar wall (x200), (E) marked thickening of the inter-lobular septa with few inflammatory cell infiltration (arrow), notice also, accumulation of inflammatory cells in pulmonary alveoli (asterisk, x200), (F) marked fibrinous thickening of the pleura with mononuclear leukocytic cellular infiltration (arrow, x100, insert, x400). Meanwhile, hyperplasia of the biliary epithelium with peri-ductal fibrosis and newly formed bile ductules (Fig. 4A) as well as peri-ductal leukocytic cellular infiltrations were observed. Additionally, severe intracytoplasmic vacuolation in the hepatocytes with congestion and dilatation of the blood sinusoids as well as multiple focal areas of coagulative necrosis were detected in the hepatic parenchyma (Fig. 4B,C). Interestingly, extensive hemorrhage in the red pulp of spleen (Fig. 4D) with marked lymphoid depletion and hyalinization of the wall of central arterioles (Fig. 4E) was found. Marked fibrinous pericarditis (Fig. 5A), congestion of myocardial blood vessels with inter-muscular leukocytic cellular infiltration (Fig. 5B) was the most common detected pathological changes in the heart. Hemorrhages and necrotic changes were demonstrated in the glomerular tuft with widening of Bowman’s space (Fig. 5C) and renal tubules. In contrast, hyper-cellularity of glomerular tuft with adhesion between glomerular tuft and Bowman’s capsule (Fig. 5D) as well as degeneration in the wall of renal blood vessels was seen in other examined cases. Meanwhile, the microscopical examination of the intestine revealed degeneration and detachment of the lining epithelium of the intestinal villi with extensive necrosis in association with congestion of submucosal blood vessels with few leucocytes infiltration (Fig. 5E).
Fig. 4. H&E stained section of liver (A-C) and spleen (D-E) obtained from goat naturally infected with P. multocida, showing (A) hyperplasia of the biliary epithelium (arrow) with peri-ductal fibrosis (asterisk, x400), (B) severe intracytoplasmic vacuolation in the hepatocytes (x 200), (C) focal areas of coagulative necrosis was detected in the hepatic parenchyma (arrow, x400), (D) extensive hemorrhage in the red pulp of spleen (x200), (E) marked lymphoid depilation and hyalinization of the wall of central arterioles (x400).
Fig. 5. H&E stained section of heart (a-b), kidney (C-D) and intestine (E) obtained from goat naturally infected with P. multocida, showing (a) fibrinous pericarditis (asterisk, x100, insert, x400), (B) inter-muscular leukocytic cellular infiltration (arrow, x 400), (C) necrotic changes in the glomerular tuft with widening of Bowman’s space (arrow, x400), (D) hyper-cellularity of glomerular tuft with adhesion between glomerular tuft and Bowman’s capsule (arrow, x400), (E) extensive necrosis of the intestinal villi (arrow) with congestion of submucosal blood vessels (C) and few leucocytes infiltration (x400). Immunohistochemical staining Positive brown IHC reaction of P. multocida antigen was demonstrated in several tissues (lung, heart, liver, kidney, spleen and intestine) (Table 1). The reaction of immunohistochemistry ranged from weak, moderate to strong. Strong and precise IHC reaction associated with p. multocida were detected Most often, in alveolar exudate, pulmonary interstitial tissue (Fig. 6A) and pulmonary blood vessels as well as in the pericardium, but moderate positive reaction was observed in the hepatic parenchyma (Fig. 6B), glomeruli and renal tubules (Fig. 6C) as well as in splenic tissue (Fig. 6D) with weak positive reaction in the intestinal mucosa. The antigen distribution normally associated strongly with lesion incidence. Interestingly, there was an association between the PCR and IHC techniques and there were samples which tested positive for the detection of P. multocida were positive by both techniques and the rate of agreement between these methods was shown in Table 1.
Fig. 6. Positive immunohistochemical reaction of P. multocida in lung, x200 (A), liver, x100 (B), kidney, x400 (C) and spleen, x400 (D) Table 1. Bacterial isolation and nucleic acid of Pasteurella multocida in tissue samples detection by PCR and IHC.
|
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Discussion |
|||||||||||||||||||||||||||||||||||
|
P. multocida causes economically significant diseases in a broad range of animal species, as it is commensal bacteria in the respiratory tract of most mammals and birds. It has been found that, P. multocida create a complex interaction between host tissues to grow rapidly and produce its pathological effect at favorable condition resulting in great economic losses (Boyce and Adler, 2006). Thus, the existing work was aimed to diagnose goat pasteurellosis via post-mortem and microscopical examination as well as PCR and immunohistochemical detection of the bacterial genome on the host tissues. In the current study, pneumonic lungs with dark red patches and consolidation of many lobes with firm consistency was the most common gross lesions observed in the lungs of most cases as P. multocida is commonly found in the upper respiratory tract of goats causing pneumonia that characterized by consolidation in the whole of the apical lobe of the pneumonic lung with bacterial infections, which supports the findings of the present study (Emikpe and Akpavie, 2010). The histopathological examination revealed varying degree of bronchopneumonia was demonstrated in different examined animals. These findings could be attributed to varying susceptibility among different infected goats. The fibrino-suppurative and necrotic inflammatory reaction are characteristic features for acute pulmonary infections due to P. multocida. The pulmonary tissue contains compact infiltrates of neutrophils, fibrin, sero-proteinaceous material and erythrocytes as well as extensive parenchymal necrosis. These histopathological changes may be due to P. multocida endotoxins and toxic proteins such as leukotoxin, lipopolysaccharide and polysaccharide (Hodgson, 2006) and also due to the inflammatory factors produced by neutrophils and other inflammatory cells (Slocombe et al., 1985). Additionally, leukotoxin has been found to be important for pathogenesis of P. multocida since it is inherently toxic to leukocytes, leading to fibrin deposition on pulmonary tissue and pleural surfaces that could clarify the occurrence of fibrinous pleuritis and pneumonia in the current study. Interestingly, the pulmonary tissues especially pulmonary alveoli and bronchioles was infiltrated with different inflammatory cells, this could be induced by the release of pro-inflammatory cytokines such as TNF-α, IL-β and IL-8 that produced in reaction to components of bacterial cell wall in alveolar air space (Locksley et al., 2001). These cytokines enhance leukocytic cellular infiltrations at the infection site (Yoshi et al., 2001). Additionally, it has been found that, presence of neutrophils in the exudate in the pulmonary tissue in the current study is due to the role of neutrophils in clearance of P. multocida from the pulmonary tissue as they are a part of innate immune response that represented mainly in both phagocytosis and production of soluble factors as complement proteins, acute phase proteins and cytokines (Muller and Kohler 1997, Thacker, 2006a). Furthermore, presence of macrophage with neutrophils mediates phagocytosis and cytokine production (Thacker, 2006b). Meanwhile, the presence of Inflammatory exudates in the bronchioles may occur due to secondary bacterial spread across respiratory tracts in the lung tissue (Praveena et al., 2010). Multifocal coagulative necrosis has been proposed to be a characteristic pulmonary lesion with a close relation to P. multocida antigens (Slocombe et al., 1985). However, in the present study Several pneumonic lungs had a fatal fibrinous pneumonia. Furthermore, combination of exudative and proliferative pneumonic lesions was observed in some cases and the pattern of interstitial pneumonia was demonstrated in few cases. Such proliferative lesions may have taken place during the repair of bronchopneumonia caused by bacteria. In the current study, thrombosis of blood vessels with detection of bacteria in the blood vessels was noticed. These findings could clarify the detection of pathological alterations in other organs such as liver, heart kidney, spleen and intestine that may be due to the ability of invasive bacteria to be taken systemically and resulted in septicemia (Emikpe and Akpavie, 2010). In the present work, P. multocida were isolated from lung, liver, heart, kidney, spleen and intestine. These findings agreed with the obtained results recorded by Sunder and Kumar (2001) who isolated P. multocida from internal organs of freshly dead Indian goats. Immunoperoxidase staining method is a viable method to detect Pasteurella sp. in paraffin embedded tissue sections (Haziroglu, et al, 1996). Remarkably, in the current research, an extensive invasion of the lung by P. multocida and moderate invasion to other organs (liver, heart, kidney and spleen) while weak reaction was observed in the intestine using IHC. However, the low concentration of bacteria in the examined tissues may result in a weak reaction in the IHC process (Haziroglu et al., 1996). The obtained results revealed that there is a correlation between the bacteriological, PCR and IHC outcomes as PCR and IHC were also positive for all culture positive samples. However, the positive rate of the bacterial antigens demonstration was higher in PCR and IHC investigated samples than the bacteriologically observed rate. These results could be due to that the live and dead bacterial antigen could be detected by PCR and IHC techniques, in contrast there is no identification of dead organisms by bacteriological cultures (Haziroglu et al., 1996). In the current work, p. multocida antigens were noticed in different organs, however, the obtained PCR results were different from IHC results. This could be explained by the uneven distribution of the bacteria within different investigated organs especially in organs that have only few bacteria. It has to be noted that immunohistochemistry labelling positive reaction for pasteurella. Additionally, P. multocida antigens localization typically had a strong correlation with presence of lesions. Accordingly, specific diffuse reactions against p. multocida likely reflect the bacterial solubilized antigen resulting from degeneration and destruction of the bacteria (Haritani et al., 1989). As a result, p. multocida antigen detection by IHC seems to be more effective than bacterial and PCR detection in various examined tissues. Although, pasturellosis in goat produce pneumonic pasteurellosis but it nevertheless contributes the incidence of the disease in the field. In the current research, the existence of pasteurella antigens in naturally infected goat pneumonia was proved using IHC, PCR and bacteriological isolation. The obtained results indicated that IHC using polyclonal antibodies can be a beneficial tool to diagnose pasteurellosis. |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Conclusion |
|||||||||||||||||||||||||||||||||||
|
The post-mortem and microscopical examination can be useful in the initial diagnosis of Pasteurellosis but needs to be enhanced by other immunodiagnostic tests. However, IHC technique proved to be a specific, reliable and rapid technique to supplement these conventional methods for diagnosis of pasteurellosis. |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Conflict of Interests |
|||||||||||||||||||||||||||||||||||
|
The author declares that no competing interests exist. |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
References |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||
|
Ackermann, M.R., Brogden, K.A., 2000. Response of the ruminant respiratory tract to Mannheimia (Pasteurella) haemolytica. Microbes. Infect. 2, 1079-1088. Bancroft, J.D., Gamble, M., 2007. Theory and Practice of Histological Techniques. fifth Ed; Churchill Livingstone, London, UK. pp. 125-138. Boyce, J.D., Adler, B., 2006. How does Pasteurella multocida respond to the host environment? Curr. Opin. Microbiol. 9, 117-122. Dabo, S.M. Taylor, J.D., Confer, A.W., 2007. Pasteurella multocida and bovine respiratory disease. Anim. Health Res. Rev. 8, 129-150. Emikpe, B.O., Akaptvie, S.O., 2010. The clinical and pathological features of experimental Mannheimia hemolytica A2 infection in ''''est African dwarf goats. Bull. Anim. Health. Prod. Afr. 58, 248-255. Gautam, R., Kumar, A.A., Singh, V.P., Dutta, T.K., Shivchandra, S.B., 2004. Specific identification of Pasteurella multocida serogroup-A isolates by PCR. Res. Vet. Sci. 76, 179-185. Haziroglu, R., Diker, K.S., Turkarslan, J., Gulbahar, M.Y., 1996. Detection of Mycoplasma ovipneumoniae and Pasteurella haemolytica Antigens by an Immunoperoxidase Technique in Pneumonic Ovine Lungs. Vet. Pathol. 33, 74-76. Haritani, M., Ishino, S., Oka, M., Nakazawa, M., Kobayashi, M., Narita, M., Takizawa, T., 1989. Immunoperoxidase evaluation of pneumonic lesions in calves naturally infected with Pasteurella haemolytica. Nippon Juigaku Zasshi 51, 1137-41. Hodgson, J.C., 2006. Endotoxin and mammalian host responses during experimental disease. J. Comp. Pathol. 135, 157-175. Locksley, R.M., Killen, N., Lenardo, M.J., 2001. The TNF and TNF receptor superfamilies-integrating: mammalian biology. Cell. 104, 487-501. Quinn, P.J., Markey, B.K., Leonard, F.C., Fitzpatrick, E.S., Fanning, S., Hartigen, P.J., 2011. Pasteurella species, Mannheimia haemolytica and Bibersteinia trehalosi. In: Ibid. (Eds), Veterinary Microbiology and Microbial Disease. second Ed. Wiley-Blackwell, Ames, Iowa, pp.300-308. Muller, G., Kohler, H., 1997. Experimental affecting of pulmonary clearance of Pasteurella multocida induced pneumonia in swine. Berl. Munch. Tierarztl. Wochenschr. 110, 378-380. Narita, M., Kimura, K., Tanimura, N., Arai, S., Katsuda, T.K., 2000. Immunohistochemical characterization of calf pneumonia produced by the combined endobronchial administration of bovine herpesvirus 1 and Pasteurella haemolytica. J. Comp. Pathol. 123, 126–34. Ozyildiz, Z., Tel, O.Y., Yilmaz, R., Ozsoy, S.Y., Keskin, O., 2013. Pathological and microbiological investigations of pneumonic pasteurellosis in sheep. J. Fac. Vet. Med. Univ. Kafkas. 19, 103-108. Praveena, P.E., Periasamy, S., Kumar, A.A., Singh, N., 2010. Cytokine profile, apoptosis and pathology of experimental Pasteurella multocida serotype A1 infection in mice. Res. Vet. Sci. 89, 332-339. Slocombe, R.F., Malark, J., Ingersoll, R., Derksen, F.J., Robinson, N.E., 1985. Importance of neutrophils in the pathogenesis of acute pneumonic pasteurellosis in calves. Am. J. Vet. Res. 46, 2253-2258. Sunder, J., Kumar, A.A., 2001. Studies on toxigenic strains of Pasteurella multocida of goat origin Indian. Vet. J., 78: 184-188. Thacker, E.L., 2006a. Lung inflammatory responses. Vet. Research. 37, 469-486. Thacker, E.L., 2006b. Mycoplasmal Disease. In: Diseases of Swine. 9th ed. Iowa State University Press, Ames, pp. 701-717. Watson, P.J., Davis, R.L., 2002. Outbreak of Pasteurella multocida septicaemia in neonatal lambs. Vet. Record. 151, 420-422. Yoshie, O., Imai, T., Nomiyana, H., 2001. Chemokines in immunity. Advances in Immunol. 78, 57-110. |
|||||||||||||||||||||||||||||||||||
|
|